
Leído para Ud.: Una horma para el zapato darwinista (3-4)

Sitios de unión: más allá del límite
La vida es mucho más que un par de mutaciones en éste o aquel gen (el cual tiene que pre-existir para que tales mutaciones ocurran en él). Behe nota que la vida depende de muchos complejos proteínicos en los cuales múltiples proteínas se asocian entre sí de manera muy específica en los llamados sitios de unión[1].![]() Por ejemplo la hemoglobina necesita cuatro proteínas para ensamblarse correctamente; si este proceso de unión falla el resultado de tal error genético es la ya mencionada anemia talasemia. Es por eso que existen proteínas chaperonas (vgr. la que ayuda en la formación de la hemoglobina se llama AHSP=proteína estabilizante alfa hemoglobina). Si la proteína chaperona no hace sus deberes habrá daño en los glóbulos rojos. Otras proteínas necesitan seis o más ensamblajes en sitios de unión específicos. Behe demuestra con lujo de detalles que:
Por ejemplo la hemoglobina necesita cuatro proteínas para ensamblarse correctamente; si este proceso de unión falla el resultado de tal error genético es la ya mencionada anemia talasemia. Es por eso que existen proteínas chaperonas (vgr. la que ayuda en la formación de la hemoglobina se llama AHSP=proteína estabilizante alfa hemoglobina). Si la proteína chaperona no hace sus deberes habrá daño en los glóbulos rojos. Otras proteínas necesitan seis o más ensamblajes en sitios de unión específicos. Behe demuestra con lujo de detalles que:
“Generar un nuevo sitio de unión entre proteínas presenta una dificultad del mismo orden o peor que el desarrollo de la resistencia a la cloroquina en el parásito de la malaria (i.e. una chance en cien trillones =1/1020). …la gran mayoría de las proteínas en la célula funcionan con complejos con seis o más sitios de unión. Totalmente más allá del límite [de la evolución]”[2].
No resulta una sorpresa entonces que no haya en la práctica ninguna nueva interacción entre proteínas surgida durante la guerra entre el parásito de la malaria y los seres humanos, ni en aquel ni en estos. Una célula típica tiene unos 10 mil sitios de unión de proteínas. De todos los sitos de unión entre proteínas estudiados solo uno ha surgido debido a mutaciones aleatorias: la condición que resulta en anemia falciforme[3]. Además de ser no-específica, destruye la estructura normal de la hemoglobina causando enfermedad, no ventaja evolucionaria.
Resumiendo, complejos proteínicos con más de dos sitios de unión diferentes –que requieren tres o más tipos de proteínas diferentes– están más allá del límite evolucionista darwiniano. Más allá de lo que es razonablemente biológico de alcanzar desde que la vida apareció en la tierra[4].
VIH-SIDA: plus ça change, plus c’est la même chose
El virus de la inmunodeficiencia humana (VIH) que causa el síndrome de inmunodeficiencia adquirida (SIDA) es de particular interés porque muta a velocidades evolucionarias records, 10 mil veces más rápido que el parásito de la malaria plasmodium. Se estima que el VIH ha producido cien trillones (1020) de copias en las últimas décadas. A pesar de esta notoria fecundidad sus bases genéticas han cambiado muy poco. “Todas y cada una de las posibles mutaciones singulares ocurre entre 10 mil y 100 mil veces por día en una persona infectada con VIH.” Y sin embargo esto no ha generado ningún “nuevo sistema o maquinaria básica”[5]. “Ninguna duplicación de genes ha sucedido que conlleve a una nueva función. Ninguno de los elaborados trucos que rutinariamente son mencionados en las especulaciones darwinistas le ha ayudado mucho al VIH”[6].
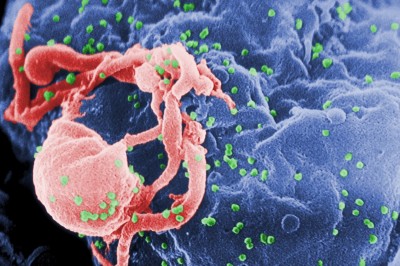
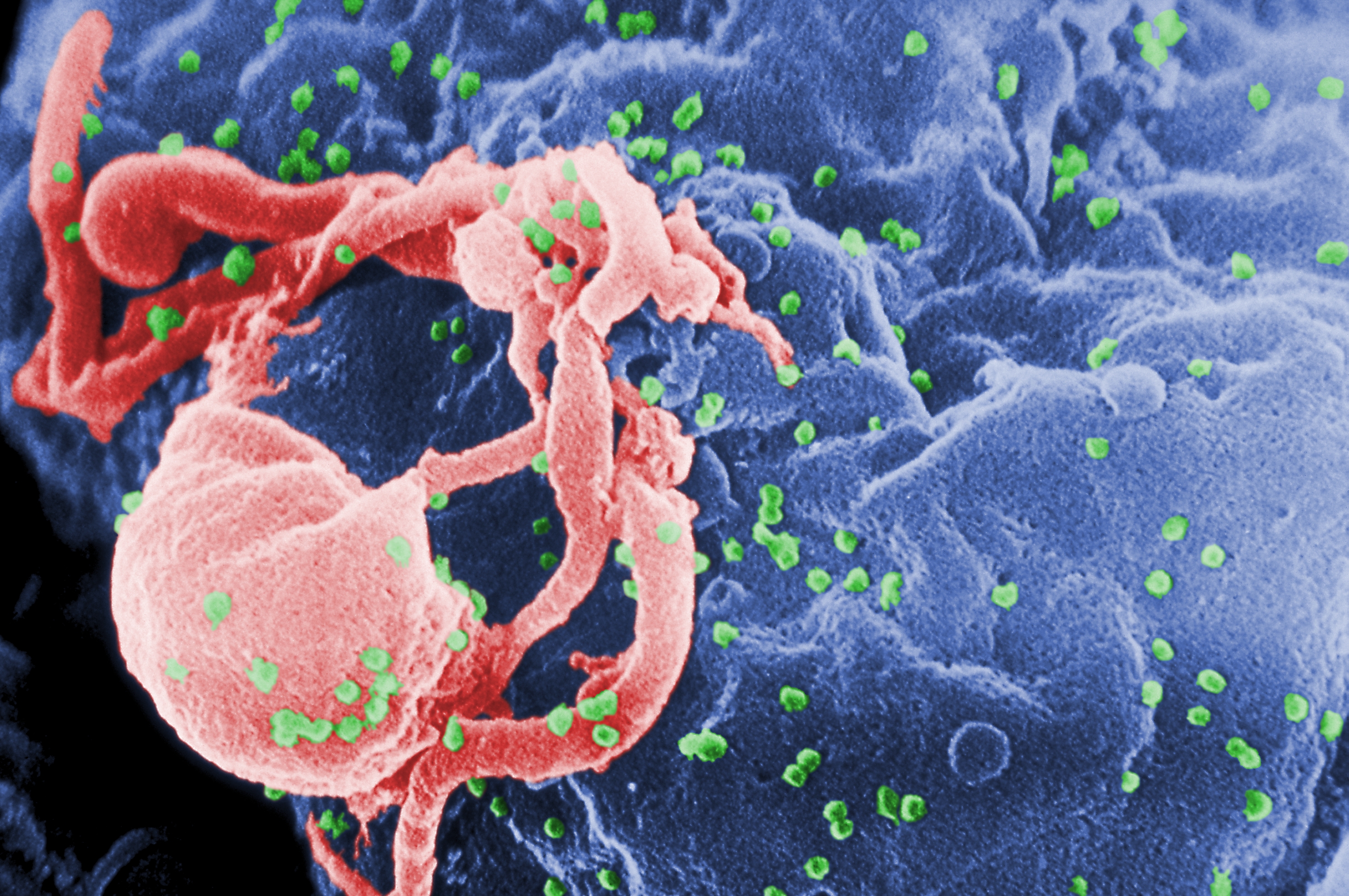
Figura 5: Imagen de microscopio electrónico con virus VIH (coloreados en verde) atacando una célula linfática. Fuente Wikimedia.
{kind=link}
Los cambios bioquímicos provechosos para sobrevivir a las drogas observados tanto en plasmodium como en VIH, han sido triviales; como por ejemplo mutaciones singulares que ligeramente cambian la forma de una enzima.
Los evolucionistas han celebrado con bombos y platillos los modestos cambios de forma y tamaño en el pico de los canarios debido al cambio de alimentación. Algo totalmente trivial en términos evolutivos. Sin embargo hay estudios genéticos sobre miles de miles de generaciones, billones de billones de organismos, que muestran que prácticamente nada bioquímicamente importante ha ocurrido.
La cantidad de parásitos plasmodium, de VIH y de E. coli (tres tipos de organismos muy diferentes entre sí en complejidad, ciclo de vida, y tipo de dominio: eukaryota, virus, prokaryota -ver Figura 1) en los últimos 50 años excede considerablemente el número total de mamíferos desde su supuesto origen evolucionario (varios cientos millones de años atrás), y sin embargo poco y nada ha sido logrado mediante evolución darwinista. Esto sugiere que los mamíferos no habrían podido generar casi ningún cambio en su marco de tiempo:
“Nuestra experiencia con el VIH da razón a pensar que el darwinismo no es capaz de lograr mucho –aun con miles de millones de años y todas las células en el mundo a su disposición”[7].
¿Son los cálculos de VIH y malaria representativos para todos los organismos? Si lo son. A nivel bioquímico hay una gran similitud en todos los organismos vivos. Con algunas pocas excepciones el código genético es el mismo para todas las millones de especies; las proteínas están hechas de los mismos aminoácidos, los ácidos nucleicos (ej. ADN) están hechos de los mismos tipos de nucleótidos. Las fuerzas físicas entre proteínas no varían de organismo a organismo, ni tampoco la forma de la proteína depende de la especie[8].
Un par de desengaños más
El descubrimiento que ciertos genes comunes, conocidos como genes Hox, controlan el desarrollo embrionario (i.e. los planos del cuerpo) de muchos organismos multicelulares fue recibido con bombos y platillos por los evolucionistas. Genes muy similares controlan la posición de los ojos tanto en insectos como en vertebrados, aunque los tipos de ojo son muy diferentes. El orden de los genes Hox se corresponde con el la estructura del cuerpo del animal.

Figura 6: Posición de los genes Hox a lo largo del cromosoma y la correspondiente zona del cuerpo de la mosca de la fruta en donde se expresan. Fuente Wikimedia.
{kind=link}
Los evolucionistas pensaron que con algún que otro toqueteo (mutaciones) los genes Hox podrían generar nuevos planes maestros de cuerpos y más aún explicar el origen de las principales categorías de vida. La excitación inicial dio lugar a estipulaciones más modestas. Las mutaciones en estos genes no generan nada fundamentalmente nuevo. Solo reacomodan lo que ya existe –por ejemplo poniendo los ojos o antenas de una mosca donde normalmente no estarían. Esto ha resultado en un callejón sin salida en términos evolutivos.
Los genes maestros de regulación son simplemente interruptores, que prenden y apagan la maquinaria pesada molecular realmente encargada de construir los órganos[9]. No explican en absoluto cómo algo fue construido.
Las hipótesis de “auto-organización” también son presa fácil para el ojo avizor de Behe. No hay ninguna evidencia que las cosas se instauren por sí mismas por defecto. No hay evidencia alguna de una capacidad innata en los organismos vivos para crear complejidades nuevas. No hay evidencia tampoco de que la célula tenga habilidad de crear nuevas funciones mediante una suerte de auto-ingeniería genética[10].
Sistemas de control, filos y clases: más allá del límite
Behe también apunta que el descubrimiento de sistemas de control –como los descriptos antes involucrados en la construcción y reparación del cilio, hace las cosas más difíciles aun para el darwinismo. Ahora la evolución tiene que explicar no sólo el origen de los genes que codifican las proteínas sino también su sistema de control:
“Los sistemas de control adosan una capa extra de complejidad por encima de la complejidad misma de las estructuras moleculares existentes –que en nuestra inocencia no habíamos imaginado que serían requeridos. La necesidad de sistemas de control no hace la tarea darwinista más fácil; la hace mucho más difícil” [11].
El problema de la evolución de los sistemas de control cobra mayor vuelo con el descubrimiento de las llamadas redes de regulación génicas (RRG) que definen la secuencia de pasos necesarios para construir los componentes del cuerpo de un animal. Las RRG son mapas lógicos que se asemejan a los usados en el diseño de circuitos electrónicos de computadoras[12] involucrando docenas de proteínas. La falla de una implica la falla del todo –ergo irreduciblemente complejos. Cada módulo de estas RRG se denomina kernel. Ahora un dato importante: cada filo (ver Figura 1) difiere en sus mapas corporales, de modo que deben tener diferentes kernels, que no pueden haber evolucionado. Ergo, sostiene Behe, el origen de cada filo en la categoría taxonómica está más allá de la frontera de la evolución[13].
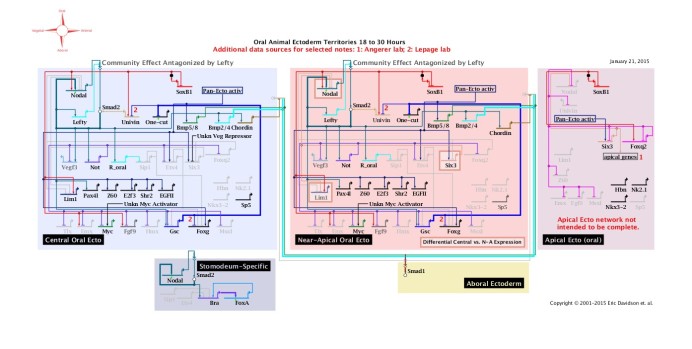
Figura 7: Esquema de una red de regulación génica. La red evoca fuertemente un complejo sistema electrónico o circuito eléctrico. Fuente Davdison Lab.
Ciertos tipos de células como las llamadas células B de nuestro sistema inmunológico están controladas por redes de regulación génicas (RRG). Debido a que diferentes clases (ver Figura 1) de vertebrados difieren en el número de diversos tipos de células que producen (e.g. los anfibios alrededor de 150, los pájaros 200, los mamíferos 250) esto es una fuerte indicación que el origen de cada clase en la categoría taxonómica está más allá de la frontera de la evolución. A esta altura Behe comienza a delinear uno de sus principales conclusiones:
“Concluimos que el diseño animal probablemente se extiende al menos hasta las clases de vertebrados, quizás más profundo aun; y que las mutaciones al azar posiblemente explica diferencias a menos hasta las especies, quizás un poco más allá. En algún lugar entre las especies y las clases vertebradas yace la frontera de la evolución darwiniana”[14].
Es decir que la evolución darwinista no puede explicar la categoría taxonómica de clase y todas aquellas por encima de ésta. Lo que significa que las principales clases de vertebrados –mamíferos, reptiles, peces, anfibios, aves– no pueden haber evolucionado al azar (ver Figura 1).
¿Se extiende el diseño a niveles de vida más profundos aun, en las categorías de órdenes y familias, diferenciando creaturas como jirafas, ballenas y murciélagos? Behe piensa que sí, pero es en éste punto que el actual conocimiento molecular confiable se acaba y prefiere no emitir juicio hasta que aparezca más información sólida[15].
Ajuste fino
En el extremo superior de la Figura 1 tenemos las más altas leyes del universo. La ciencia ha revelado un sorprendente grado de ajuste de estas leyes con el fin de favorecer la vida en la tierra. Lo que le da al dicho del Martin Fierro “hacéte amigo del juez” una perspectiva más alta. Varios físicos han observado este notorio ajuste[16].
Las propiedades de las partículas físicas y elementos químicos también están ajustadas para facilitar la vida en la tierra. El agua sigue siendo un caso ejemplar: cuando todos los líquidos se contraen al congelarse, el agua se expande, un dato para nada trivial que dejamos al lector explorar[17].
Ciertos detalles también están ajustados para favorecer la vida en la tierra. Un planeta tiene que estar en la región adecuada del sistema solar para evitar cocinarse o congelarse; y en la región adecuada de la galaxia para evitar ser calcinado por altas dosis de rayos X o quedar desprotegido por falta de un campo magnético[18].
Este ajuste fino que promueve la vida en la tierra abarca desde las más básicas leyes físicas hasta el mismo tejido de la vida, pasando por el origen mismo de la vida[19].